|
THE AGES OF GAIA: A BIOGRAPHY OF OUR LIVING EARTH |
||||
|
2: What Is Gaia?
Travel back in your memory to the time when you first awoke, that exquisite moment of childhood when first you came alive -- the sudden rush of sound and sight, as if a television receiver had been switched on and was about to bring news of vast importance. I seem to recall sunlight and soft fresh air; then suddenly knowing who I was and how good it was to be alive. To reminisce about the first memory of my personal life may seem irrelevant in our quest to understand Gaia. But it isn't. As a scientist I observe, measure, analyze, and describe phenomena. Before I can do these things I need to know what I am observing. In a broad sense it may be unnecessary to recognize a phenomena when observing it, but scientists almost always have preconceived notions of the object of their study. As a child I recognized life intuitively. As an adult wondering about the Earth's strange atmosphere -- a mixture made of incompatible gases such as oxygen and methane coexisting like foxes and rabbits in the same burrow -- I was forced to recognize Gaia, to intuit her existence, long before I could describe her in proper scientific terms. The concept of Gaia is entirely linked with the concept of life. To understand what Gaia is, therefore, I first need to explore that difficult concept, life. They hate to admit it, but the life scientists, whether the natural historians of the nineteenth century or the biologists of the twentieth, cannot explain what life is in scientific terms. They all know what it is, as we have done since childhood; but in my view no one has yet succeeded in defining life. The idea of life, the sense of being alive, are the most familiar and the most difficult to understand of the concepts we meet. I have long thought that the answer to the question "What is life?" was deemed so important to our survival that it was classified "top secret" and kept locked up as an instinct in the automatic levels of the mind. During evolution, there was great selection pressure for immediate action: crucial to our survival is the instant distinction of predator from prey and kin from foe, and the recognition of a potential mate. We cannot afford the delay of conscious thought or debate in the committees of the mind. We must compute the imperatives of recognition at the fastest speed and, therefore, in the earliest-evolved and unconscious recesses of the mind. This is why we all know intuitively what life is. It is edible, lovable, or lethal. Life as an object of scientific inquiry requiring precise definition is much more difficult. Even scientists, who are notorious for their indecent curiosity, shy away from defining life. All branches of formal biological science seem to avoid the question. In the Dictionary of Biology compiled by M. Abercrombie, C. J. Hickman, and M. L. Johnson, these three distinguished biologists succinctly define all manner of words like ontogeny (development), Pteridophyta (ferns), and ecdysis (a stage in insect development). Under the letter L there is leptotene (the first sign of chromosome pairing in meiosis) and limnology (the study of lakes), but nowhere is life mentioned. When the word life does appear in biology it is in rejection, as by the philosophically inclined N. W. Pirie who, in 1937, published an article entitled "The Meaninglessness of the Terms 'Life' and 'Living'." The Webster and the Oxford dictionaries are not much more help. Both remind of the word's origin from the Anglo-Saxon lif. This may explain some of the reluctance of academic biologists to tangle with so elemental a concept as life. The tribal war between the Normans and the Saxons was long enduring; the medieval schoolmen, knowing where power and preference lay, chose to support the victorious Norman establishment and to keep Latin as their language. Life was another of those rude uncivilized Anglo-Saxon words, best avoided in polite company. The Latin equivalent of lif, anima, was even less help. It was close in meaning to that other four-letter Gothic word, soul. To go back to the Webster dictionary, it defines life as:
The Oxford dictionary says much the same:
If such manifestly inadequate definitions of life are all I have to work with, can I do much better defining the living organism of Gaia? I have found it very difficult, but if I am to tell you about it I must try. I can start with some simpler definitions and classifications. Living things such as trees and horses and even bacteria can easily be perceived and recognized because they are bounded by walls, membranes, skin, or waxy coverings. Using energy directly from the Sun and indirectly from food, living systems incessantly act to maintain their identity, their integrity. Even as they grow and change, grow and reproduce, we do not lose track of them as visible, recognizable entities. Although there are uncountable millions of individual organisms all growing and changing, their traits in common allow us to group them and recognize that they belong to species such as peacocks, dogs, or wheat. About ten million species are estimated to exist. When any individual fails to get energy and food, fails to act to maintain its identity, we realize it is moribund or dead. An important step in our understanding is to recognize the significance of collections of living things. You and I are both composed of a collection of organs and tissues. The many beneficiaries of heart, liver, and kidney transplants testify eloquently that each of these organs can exist independently of the body when kept warm and supplied with nutrients. The organs themselves are made up of billions of living cells, each of which can also live independently. Then the cells themselves, as Lynn Margulis has shown, are communities of microorganisms that once lived free. The energy-transforming entities of animal cells (the mitochondria) and of plants (the mitochondria and the chloroplasts) both were once bacteria living independently. Life is social. It exists in communities and collectives. There is a useful word in physics to describe the properties of collections: colligative. It is needed because there is no way to express or measure the temperature or the pressure of a single molecule. Temperature and pressure, say the physicists, are the colligative properties of a sensible collection of molecules. All collections of living things show properties unexpected from a knowledge of a single one of them. We, and some other animals, keep a constant temperature whatever the temperature of our surroundings. This fact could never have been deduced from the observations of a single cell from a human being. The tendency to constancy was first noted by the French physiologist Claude Bernard in the nineteenth century. His American successor in this century, Walter Cannon, called it homeostasis or the wisdom of the body. Homeostasis is a colligative property of life.
We have no trouble with the idea that noble entities such as people are made up from an intricate interconnected set of cell communities. We don't find it too difficult to consider a nation or a tribe as an entity made up of its people and the territory they occupy. But what of large entities, like ecosystems and Gaia? It took the view of the Earth from space, either directly through the eyes of an astronaut, or vicariously through the visual media, to give us the personal sense of a real live planet on which the living things, the air, the oceans, and the rocks all combine in one as Gaia. The name of the living planet, Gaia, is not a synonym for the biosphere. The biosphere is defined as that part of the Earth where living things normally exist. Still less is Gaia the same as the biota, which is simply the collection of all individual living organisms. The biota and the biosphere taken together form part but not all of Gaia. Just as the shell is part of a snail, so the rocks, the air, and the oceans are part of Gaia. Gaia, as we shall see, has continuity with the past back to the origins of life, and extends into the future as long as life persists. Gaia, as a total planetary being, has properties that are not necessarily discernible by just knowing individual species or populations of organisms living together. The Gaia hypothesis, when we introduced it in the 1970s, supposed that the atmosphere, the oceans, the climate, and the crust of the Earth are regulated at a state comfortable for life because of the behavior of living organisms. Specifically, the Gaia hypothesis said that the temperature, oxidation state, acidity, and certain aspects of the rocks and waters are at any time kept constant, and that this homeostasis is maintained by active feedback processes operated automatically and unconsciously by the biota. Solar energy sustains comfortable conditions for life. The conditions are only constant in the short term and evolve in synchrony with the changing needs of the biota as it evolves. Life and its environment are so closely coupled that evolution concerns Gaia, not the organisms or the environment taken separately. Most of my working life has been spent on the fringes of the life sciences, but I do not think of myself a biologist, nor do I believe would biologists accept me as one of them. When seen from outside, much of biology appears to be the building of data bases -- making the "whole life catalog." Sometimes, in a pensive mood, I fancy that to biologists the living world is a vast set of book collections held in interconnected libraries. In this dream, the biologists are like competent librarians who devise the most intricate classification of every new library they discover but never read the books. They sense that something is missing from their lives, and this feeling intensifies as new collections of books grow hard to find. I see the biologists expressing an almost palpable sense of relief when joined by molecular biologists who dare to start the even greater task of classifying the words the books contain. It means that the search for the answer to the awesome question of what the books are about can be put off until the new and infinitely detailed molecular classification is complete. My imaginary world, populated by biologists as book collectors, is in no way intended as a slur on the life sciences. Left to my own devices in such a world I should have been much less constructive. Impatient of waiting for an answer to the question, "What is the meaning of the books?" I would have seized some of them for experimental tests -- for example, burning them in a calorimeter and measuring, accurately, the heat released. My sense of frustration would not have lessened when I discovered that the densely packed pages of an encyclopedia give no more heat than the same mass of plain paper. Like the biologists' classification, this physical experiment would have been profoundly unsatisfying because it would have put to Nature the wrong question. Can we scientists, any of us, do better in our quest to understand life? There are three equally powerful approaches: molecular biology, the understanding of those information-processing chemicals that are the genetic basis of all life on Earth; physiology, the science concerned with living systems seen holistically; thermodynamics, the branch of physics that deals with time and energy and that connects living processes to the fundamental laws of the Universe. Of these sciences, the latter is the one that may go furthest in the quest to define life, yet so far has made the least progress. Thermodynamics grew from down-to-earth origins, the quest of engineers to make steam engines more efficient. It flourished in the last century, both taxing and entertaining the minds of the greatest scientists. The first law of thermodynamics is about energy, or in other words, the capacity to do work. Energy, says the first law, is conserved. Energy in the form of sunlight falling on the leaves of a tree is used in many ways. Some is reflected so that we see the leaves as green, some is absorbed and warms them, and some is transformed to food and oxygen; ultimately, we eat the food, consume it with the oxygen we breathe, and so use the Sun's energy to move, to think, and to keep warm. The first law says that this energy is always conserved and that no matter how far it is dispersed the total always remains the same. The second law is about the dissymmetry of Nature. When heat is turned to work, some of it is wasted. The redistribution of the total quantity of energy in the Universe has direction, says the second law. It is always running down. Hot objects cool, but cool objects never spontaneously become hot. The law can appear to be broken when some metastable store of internal energy is tapped, as when a match is struck, or a piece of plutonium experiences nuclear fission, but once used up the energy cannot be recovered. The law was not broken, the energy was merely redistributed and the downward path maintained. Water does not flow up the rivers from the sea to the mountains. Natural processes always move towards an increase of disorder, and this disorder is measured by entropy. It is a quantity that always and inexorably increases. Entropy is real, not some hazy notion invented by professors to make it easier to challenge students with difficult examination questions. Like the length of a piece of string or the temperature of wine in a glass, it is a measurable physical quantity. Indeed, like temperature, the entropy of a substance is, in a practical sense, zero at the absolute zero of -- 273°C. When heat is added to a material substance, not only the temperature increases but also the entropy. Unfortunately there is a complication: whereas temperature can be measured with a thermometer, entropy cannot be measured directly with an "entropometer." Entropy, measured in the units calories per gram per degree, is the total quantity of heat added, divided by the temperature. Consider the lifeless perfection of a snowflake, a crystal so exquisitely ordered in its fractal pattern that it is one of the most intricate of nonliving things. The quantity of heat needed to melt a snowflake to a raindrop is 80 times larger than the quantity needed to warm the raindrop by a single degree of temperature. The increase of entropy when snowflakes melt is 80 times larger than when they warm from -1°C to the melting point. Alternatively, the formation of ice that expresses the ordered perfection of a snowflake represents a decrease of entropy of the same amount. Entropy is connected in quantitative terms with the orderliness of things. The greater the order, the lower the entropy. I like to think of entropy as the quantity that expresses the most certain property of our present Universe: its tendency to run down, to burn out. Others see it as the direction of time's arrow, a progression inevitably from birth to death. Far from being something tragic or a cause of sorrow, this universal tendency to decay benefits us. Without the decay of the Universe there could have been no Sun, and without the superabundant consumption of its energy store the Sun could never have provided the light that let us be. The second law is the most fundamental and unchallenged law of the Universe; not surprisingly, no attempt to understand life can ignore it. The first book I read on the question of life was by the Austrian physicist, Erwin Schrodinger. He was curious about biology and wondered if the behavior of the fundamental molecules of life could be explained by physics and biology. His famous little book, entitled What Is Life?, is a collection of the public lectures on this topic that he gave in Dublin during his exile there in the Second World War. He describes his objective on the first page:
He goes on to write:
In those times, physicists were accustomed to exploring the dead, near-equilibrium world of "periodic crystals" -- crystals whose regularity is predictable, one atom of one kind always following another of a different kind in a repeating pattern. Even these comparatively simple structures were complex enough to stretch to the limit the simple equipment then available. Organic chemists were discovering the intricate structures of the "aperiodic crystals" from living matter, such as the proteins, polysaccharides, and nucleic acids. They were still far from the present-day understanding of the chemical nature of genetic material. Schrodinger concluded that, metaphorically, the most amazing property and characteristic of life is its ability to move upstream against the flow of time. Life is the paradoxical contradiction to the second law, which states that everything is, always has been, and always will be running down to equilibrium and death. Yet life evolves to ever-greater complexity and is characterized by an omnipresence of improbability that would make winning a sweepstake every day for a year seem trivial by comparison. Even more remarkable, this unstable, this apparently illegal, state of life has persisted on the Earth for a sizable fraction of the age of the Universe itself. In no way does life violate the second law; it has evolved with the Earth as a tightly coupled system so as to favor survival. It is like a skilled accountant, never evading the payment of required tax but also never missing a loophole. Most of Schrodinger's book is an optimistic prediction of how life is knowable. The eminent molecular biologist, Max Perutz, has recently commented that little in Schrodinger's book is original, and what is original is often wrong. This may be true; but I, like many of my colleagues, still acknowledge a debt to Schrodinger for having set us thinking in a productive way. The great physicist Ludwig Boltzmann expressed the meaning of the second law in an equation of great seemliness and simplicity: S = k(lnP), where S is that strange quantity entropy; k is a constant rightly called the Boltzmann constant; and lnP is the natural logarithm of the probability. It means what it says -- the less probable something is, the lower is its entropy. The most improbable thing of all, life, is therefore to be associated with the lowest entropy. Schrodinger was not happy to associate something as significant as life with a diminished quantity, entropy. He proposed, instead, the term "negentropy," the reciprocal of entropy -- that is, 1 divided by entropy or 1/S. Negentropy is large, of course, for improbable things like living organisms. To describe the burgeoning life of our planet as improbable may seem odd. But imagine that some cosmic chef takes all the ingredients of the present Earth as atoms, mixes them, and lets them stand. The probability that those atoms would combine into the molecules that make up our living Earth is zero. The mixture would always react chemically to form a dead planet like Mars or Venus. Often in science the same idea is thought of in different contexts in different parts of the world. There is nothing occult about this. Ideas are in continuous use as currency in the exchanges between scientists and, like money, can be used to buy many different things. When Schrodinger was lecturing about negentropy in Dublin, Claude Shannon was investigating a similar quantity in the United States, but from a radically different perspective. Shannon, at the Bell Telephone Laboratories, was developing information theory. It started as a plain engineering quest to discover the physical factors that caused a message sent by cable or by radio to lose information as it passed from the sender to the receiver. Shannon soon discovered a quantity that always tended to increase; the size of the increase was a measure of the loss of information. In no experiment was the size of this quantity ever observed to decrease. On the advice of John Von Neumann, a mathematical physicist, Shannon named this quantity entropy because it so much resembled the entropy of the steam engineers. The reciprocal of Shannon's entropy is the quantity often called information. If we assume that the entropy Shannon discovered is the same as the entropy of the steam engineers, then the elusive quantity that Schrodinger associated with the improbability of life -- negentropy -- is comparable with Shannon's information. In mathematical terms, if S is the entropy then both negentropy and information are 1/S. The reward that comes from persevering with thoughts about these difficult concepts is insight to illuminate our quest to understand life and Gaia. The contribution from Shannon's theory is that information is not just knowledge. Information, in thermodynamic terms, is a measure of the absence of ignorance. Better to know all about a simple system than merely a great deal about a complex one. The less the ignorance, the lower the entropy. This is why it is so difficult to grasp the concept of Gaia from the voluminous but isolated knowledge of a single scientific discipline. If the second law tells us that entropy in the Universe is increasing, how does life avoid the universal tendency for decay? A physicist in Britain, J. D. Bernal, tried to balance the books. In 1951, he wrote in recondite terminology: "Life is one member of the class of phenomena which are open or continuous reaction systems able to decrease their internal entropy at the expense of free energy taken from the environment and subsequently rejected in degraded form." Many other scientists have expressed these words as a mathematical equation. Among the clearest and most readable are the statements in a small book, The Thermodynamics of the Steady State, written by a physical chemist, K. G. Denbigh. They can be restated less rigorously but more comprehensibly as follows. By the act of living, an organism continuously creates entropy and there will be an outward flux of entropy across its boundary. You, as you read these words, are creating entropy by consuming oxygen and the fats and sugars stored in your body. As you breathe, you excrete waste products high in entropy into the air, such as carbon dioxide, and your warm body emits to your surroundings infrared radiation high in entropy. If your excretion of entropy is as large or larger than your internal generation of entropy, you will continue to live and remain a miraculous, improbable, but still legal avoidance of the second law of the Universe. "Excretion of entropy" is just a fancy way of expressing the dirty words excrement and pollution. At the risk of having my membership card of the Friends of the Earth withdrawn, I say that only by pollution do we survive. We animals pollute the air with carbon dioxide, and the vegetation pollutes it with oxygen. The pollution of one is the meat of another. Gaia is more subtle, and, at least until humans appeared, polluted this region of the Solar System with no more than the gentle warmth of infrared radiation.
In recent times, some interesting insights have come from the investigations of Ilya Prigogine and his colleagues into the thermodynamics of eddies, vortices, and many other transient systems that are low in entropy. Things like eddies and whirlpools develop spontaneously when there is a sufficient flux of free energy. It was in the nineteenth century that a British physicist, Osborne Reynolds, curious about the conditions that led to turbulence in the flow of fluids, discovered that the onset of eddies in a stream or in a flow of gas takes place only when the flow exceeds a critical value. A useful analogy here is that if you blow a flute too gently no sound emerges. But if you blow hard enough, wind eddies form and are made part of the system that makes sound. Extending the earlier mathematics of the American physical chemist Lars Onsager, Prigogine and his colleagues have applied the thermodynamics of the steady state to develop what might be called the thermodynamics of the "unsteady state." They classify these phenomena by the term "dissipative structures." They have structure, but not the permanency of solids; they dissipate when the supply of energy is turned off. Living organisms include dissipative structures within them, but the class is broadly based. It includes many manufactured things, such as refrigerators, and natural phenomena such as flames, whirlpools, hurricanes, and certain peculiar chemical reactions. Living things are so infinitely complex in comparison with the dissipative structures of the fluid state that many feel that, although on the right track, present-day thermodynamics has far to go in defining life. Physicists, chemists, and biologists, although not rejecting these notions, do not make them part of the inspiration of their working lives. Their response is like that of a wealthy congregation to the exhortations of their priest on the virtues of poverty. It is something felt to be good, but not a way of life for next week. A crucial insight that comes from Schrodinger's generalizations about life is that living systems have boundaries. Living organisms are open systems in the sense that they take and excrete energy and matter. In theory, they are open as far as the bounds of the Universe; but they are also enclosed within a hierarchy of internal boundaries. As we move in towards the Earth from space, first we see the atmospheric boundary that encloses Gaia; then the borders of an ecosystem such as the forests; then the skin or bark of living animals and plants; further in are the cell membranes; and finally the nucleus of the cell and its DNA. If life is defined as a self-organizing system characterized by an actively sustained low entropy, then, viewed from outside each of these boundaries, what lies within is alive. You may find it hard to swallow the notion that anything as large and apparently inanimate as the Earth is alive. Surely, you may say, the Earth is almost wholly rock and nearly all incandescent with heat. I am indebted to Jerome Rothstein, a physicist, for his enlightenment on this, and other things. In a thoughtful paper on the living Earth concept (given at a symposium held in the summer of 1985 by the Audubon Society) he observed that the difficulty can be lessened if you let the image of a giant redwood tree enter your mind. The tree undoubtedly is alive, yet 99 percent is dead. The great tree is an ancient spire of dead wood, made of lignin and cellulose by the ancestors of the thin layer of living cells that go to constitute its bark. How like the Earth, and more so when we realize that many of the atoms of the rocks far down into the magma were once part of the ancestral life from which we all have come. When the Earth was first seen from outside and compared as a whole planet with its lifeless partners, Mars and Venus, it was impossible to ignore the sense that the Earth was a strange and beautiful anomaly. Yet this unconventional planet probably would have been kept in the scullery, like Cinderella, had not NASA in the role of Prince offered a rescue by way of the planetary exploration program. As we saw in chapter 1, the questions raised by space science were at first narrowly focused on a practical question: How is life on another planet to be recognized? Because that question could not be explained solely by conventional biology or geology, I became preoccupied with another question: What if the difference in atmospheric composition between the Earth and its neighbors Mars and Venus is a consequence of the fact that the Earth alone bears life? The least complex and most accessible part of a planet is its atmosphere. Long before the Viking spacecraft landed on Mars, or the Russian Venera landed on Venus, we knew the chemical compositions of their atmospheres. In the middle 1960s, telescopes tuned to the infrared radiation reflected by the molecules of atmospheric gases were used to view Mars and Venus. These observations revealed the identity and proportion of the gases with fair accuracy. Mars and Venus both had atmospheres dominated by carbon dioxide, with only small proportions of oxygen and nitrogen. More important, both had atmospheres close to the chemical equilibrium state; if you took a volume of air from either of those planets, heated it to incandescence in the presence of a representative sample of rocks from the surface, and then allowed it to cool slowly, there would be little or no change in composition after the experiment. The Earth, by contrast, has an atmosphere dominated by nitrogen and oxygen. A mere trace of carbon dioxide is present, far below the expectation of planetary chemistry. There are unstable gases such as nitrous oxide, and gases such as methane that react readily with the abundant oxygen. If the same heating-and-cooling experiment were tried with a sample of the air that you are now breathing, it would be changed. It would become like the atmospheres of Mars and Venus: carbon dioxide dominant, oxygen and nitrogen greatly diminished, and gases such as nitrous oxide and methane absent. It is not too far-fetched to look on the air as like the gas mixture that enters the intake of an internal combustion engine: combustible gases, hydrocarbons, and oxygen mixed. The atmospheres of Mars and Venus are like the exhaust gases, all energy spent. The amazing improbability of the Earth's atmosphere reveals negentropy and the presence of the invisible hand of life. Take for example oxygen and methane. Both are present in our atmosphere in constant quantities; yet in sunlight they react chemically to give carbon dioxide and water vapor. Anywhere you travel on the Earth's surface to measure it, the methane concentration is about 1.5 parts per million. Close to 1,000 million tons of methane must be introduced into the atmosphere annually to maintain methane at a constant level. In addition, the oxygen used in oxidizing the methane must be replaced -- at least 2,000 million tons yearly. The only feasible explanation for the persistence of this unstable atmosphere at a constant composition, and for periods vastly longer than the reaction times of its gases, is the influence of a control system, Gaia. It is often difficult to recognize the larger entity of which we are a part; as the saying goes, "You can't see the forest for the trees." So it was with the Earth itself before we shared with the astronauts vicariously that stunning and awesome vision; that impeccable sphere that punctuates the division of the past from the present. This gift, this ability to see the Earth from afar, was so revealing that it forced the novel top-down approach to planetary biology. The conventional wisdom of biology on Earth itself had always been forced to take a bottom-up approach by the sheer size of the Earth when compared with us or any living thing we knew. The two approaches are complementary. In the understanding of a microbe, an animal, or a plant, the top-down physiological view of life as a whole system harmoniously merges with the bottom-up view originating with molecular biology: that life is an assembly made from a vast set of ultramicroscopic parts. Since James Hutton there has been a "loyal opposition" of scientists who doubted the conventional wisdom that the evolution of the environment is determined by chemical and physical forces alone. Vernadsky adopted Suess's concept of the biosphere to define the boundaries of the realm of the biota. Since Vernadsky, there has been a continuous tradition (called biogeochemistry) in the Soviet Union -- and, to a lesser extent, elsewhere -- that has recognized the interaction between the soils, oceans, lakes, and rivers and the life they bear. It is well stated by a Russian, M. M. Yermolaev, in An Introduction to Physical Geography: "The biosphere is understood as being that part of the geographical envelope of the Earth, within the boundaries of which the physico-geographical conditions ensure the normal work of the enzymes." More recent members of this scientific opposition have included the following: Alfred Lotka of John Hopkins University, and Eugene Odum, who alone among ecologists took a physiological view of ecosystems; two Americans of European origin, the limnologist G. Evelyn Hutchinson and the paleontologist Heinz A. Lowenstam; an oceanographer from Britain, A. Redfield; and a Swedish geochemist, L. G. Sillen. They all have recognized the importance of the participation by life in the evolution of the environment. Most geologists, however, have neglected the presence of living organisms as an active participant in their theories of the Earth's evolution. The counterpart of this geological apartheid is the failure of most biologists to recognize that the evolution of the species is strongly coupled with the evolution of their environment. For example, in 1982 there appeared a book, Evolution Now: A Century after Darwin, edited by John Maynard Smith, which consisted of a collection of essays by distinguished biologists on the most controversial issues of evolutionary biology. In this collection, the only (and enigmatic) mention of the environment is in an essay by Stephen Jay Gould: "Organisms are not billiard balls, struck in a deterministic fashion by the cue of natural selection and rolling to optimal positions on life's table. They influence their own destiny in interesting and complex and comprehensible ways. We must put this concept of organism back into evolutionary biology." Apart from Lynn Margulis, the only other biologist I know to have taken the environment into account when considering life is J. Z. Young. In 1971, this distinguished physiologist was independently moved to write in a chapter on homeostasis in his book, An Introduction to the Study of Man: "The entity that is maintained intact, and of which we all form a part, is not the life of one of us, but in the end the whole of life upon the planet." J. Z. Young's view serves as a link between Gaia theory and the general scientific consensus. Through Gaia theory, I see the Earth and the life it bears as a system, a system that has the capacity to regulate the temperature and the composition of the Earth's surface and to keep it comfortable for living organisms. The self-regulation of the system is an active process driven by the free energy available from sunlight. The early reaction, soon after the Gaia hypothesis was introduced in the early 1970s, was ignorance in the literal sense. For the most part the Gaian idea was ignored by professional scientists. It was not until the late 1970s that it was subjected to criticism. Good criticism is like bathing in an ice-cold sea. The sudden chill of immersion in what seems at first a hostile medium soon stirs the blood and sharpens the senses. My first reaction on reading W. Ford Doolittle's criticism of the Gaia hypothesis in CoEvolution Quarterly in 1979 was shock and incoherent disbelief. The article was splendidly put together and beautifully written, but this did not lessen its frigidity. Icy waters may be pellucid, but this does not make them warm. After an icy plunge, however, comes that warm sense of relaxation when sunning on the beach. After a while, I began to realize that Ford Doolittle's criticism could be taken not so much as an attack on Gaia but as a criticism of the inadequacy of its presentation. Gaia had first been seen from space and the arguments used were from thermodynamics. To me it was obvious that the Earth was alive in the sense that it was a self-organizing and self-regulating system. To Ford Doolittle, from his world of molecular biology, it was equally obvious that evolution by natural selection could never lead to "altruism" on a global scale. He was supported in the similarly forceful and effective writings of Richard Dawkins in his book, The Extended Phenotype (1982). From their world of microscopes, how could the "selfish" interests of living cells be expressed at the distance of a planet? For these competent and dedicated biologists, positing the regulation of the atmosphere by microbial life seemed as absurd as expecting the legislation of some human government to affect the orbit of Jupiter. I am indebted to them both for having shown clearly that we were taking far too much for granted, and that Gaia lacked a firm theoretical basis. Not only did molecular biologists object to Gaia. Two other valued critics were the climatologist Stephen Schneider from Colorado, and the geochemist H. D. Holland from Harvard. They, in common with most of their peers, preferred to explain the facts of the evolution of the rocks, the ocean, the air, and the climate by chemical and physical forces alone. In his book The Chemical Evolution of the Atmosphere and the Oceans, Holland wrote: "I find the hypothesis intriguing and charming, but ultimately unsatisfactory. The geologic record seems much more in accord with the view that the organisms that are better able to compete have come to dominate, and that the Earth's near surface environment and processes have accommodated themselves to changes wrought by biological evolution. Many of these changes must have been fatal or near fatal to parts of the contemporary biota. We live on an Earth that is the best of all worlds but only for those who have adapted to it." Stephen Schneider's objection -- expressed in his book with Randi Londer, The Coevolution of Climate and Life -- was to the implication in the early papers on Gaia that homeostasis was the only means of climate regulation. I am indebted to all of these critics for having shown clearly that we were taking too much for granted, and that Gaia lacked a firm theoretical basis. Greater than this is my gratitude to Stephen Schneider who made sure that Gaia was properly debated by the scientific community by calling a Chapman Conference of the American Geophysical Union in March 1988. How in the world could the bacteria, the trees, and the animals have a conference to decide optimum conditions? How could organisms keep oxygen at 21 percent and the mean temperature at 20°C? Not seeing a mechanism for planetary control, they denied its existence as a phenomenon and branded the Gaia hypothesis as teleological. This was a final condemnation. Teleological explanations, in academe, are a sin against the holy spirit of scientific rationality; they deny the objectivity of Nature.
But when making this severest of criticisms of Gaia, the scientists may not have noticed the extent of their own errors. The innocent use of that slippery concept "adaptation" is another path to damnation. Earth is indeed the best of all worlds for those who are adapted to it. But the excellence of our planet takes on a different significance in the light of the evidence that geochemists themselves have gathered. Evidence that shows the Earth's crust, oceans, and air to be either directly the product of living things or else massively modified by their presence. Consider how the oxygen and nitrogen of the air come directly from plants and microorganisms, and how the chalk and limestone rocks are the shells of living things once floating in the sea. Life has not adapted to an inert world determined by the dead hand of chemistry and physics. We live in a world that has been built by our ancestors, ancient and modern, and which is continuously maintained by all things alive today. Organisms are adapting in a world whose material state is determined by the activities of their neighbors; this means that changing the environment is part of the game. To think otherwise would require that evolution was a game with rules like cricket or baseball -- one in which the rules forbad environmental change. If, in the real world, the activity of an organism changes its material environment to a more favorable state, and as a consequence it leaves more progeny, then both the species and the change will increase until a new stable state is reached. On a local scale adaptation is a means by which organisms can come to terms with unfavorable environments, but on a planetary scale the coupling between life and its environment is so tight that the tautologous notion of "adaptation" is squeezed from existence. The evolution of the rocks and the air and the evolution of the biota are not to be separated. It is a tribute to the success of biogeochemistry that most Earth scientists today agree that the reactive gases of the atmosphere are biological products. But most would disagree that the biota in any way control the composition of the atmosphere, or any of the important variables, such as global temperature and oxygen concentration, which depend on the atmosphere. There are two principal objections to Gaia, first that it is teleological, and that for the regulation of the climate, the chemical composition on a planetary scale, a kind of forecasting, a clairvoyance, would be needed. The second objection, most clearly expressed by Stephen Schneider, is that biological regulation is only partial, and that the real world is a "coevolution" of life and the inorganic. The second criticism is the more difficult, and in many ways the purpose of this book is to try to answer it. The first, the teleological criticism, I think is wrong and I will now try to show why. I knew that there was little point in gathering more evidence about the now-obvious capacity of the Earth to regulate its climate and composition. Mere evidence by itself could not be expected to convince mainstream scientists that the Earth was regulated by life. Scientists usually want to know how it works; they want a mechanism. What was needed was a Gaian model. In those hybrid sciences of biogeochemistry and biogeophysics, models of environmental change do not permit a regulatory role to the biota. The practitioners of these sciences assume that the operating points of the system are fixed by chemical and physical properties. For example, snow melts or forms at 0°C. The reflection of sunlight by snow cover can provide a powerful positive feedback on cooling, and a system for regulating the climate could be based on the melting or formation of snow. But there is no way for the melting point of snow, which is a characteristic of ice as a substance, to change to a more comfortable warmth of, say, 20°C. In great contrast, the operating points of a living organism are always set at favorable levels. In what way do Gaian models differ from the conventional biogeochemical ones? Does the assumption of the close coupling of life and its environment change the nature of the whole system? Is homeostasis a reasonable prediction of Gaia theory? The difficulty in answering these questions comes from the sheer complexity of the biota and the environment, and because they are interconnected in multiple ways. There is hardly a single aspect of their interaction that we can confidently describe by a mathematical equation. A drastic simplification was needed. I wrestled with the problem of reducing the complexity of life and its environment to a simple scheme that could enlighten without distorting. Daisyworld was the answer. I first described this model in 1982 at a conference on biomineralization in Amsterdam, and published a paper, "The Parable of Daisyworld," in Tellus in 1983 with my colleague Andrew Watson. I am indebted to Andrew for the clear, graphic way of expressing it in formal mathematical terms in this paper. Picture a planet about the same size as the Earth, spinning on its axis and orbiting, at the same distance as the Earth, a star of the same mass and luminosity as the Sun. This planet differs from the Earth in having more land area and less ocean, but it is well watered, and plants will grow almost anywhere on the land surfaces when the climate is right. This is the planet Daisyworld, so called because the principal plant species are daisies of different shades of color: some dark, some light, and some neutral colors in between. The star that warms and illuminates Daisyworld shares with our Sun the property of increasing its output of heat as it ages. When life started on Earth some 3.8 billion years ago, the Sun was about 30 percent less luminous than now. In a few more billion years, it will become so fiercely hot that all life that we know will die or be obliged to find another home planet. The increase of the Sun's brightness as it ages is a general and undoubted property of stars. As the star bums hydrogen (its nuclear fuel) helium accumulates. The helium, in the form of a gaseous ash, is more opaque to radiant energy than is hydrogen and so impedes the flow of heat from the nuclear furnace at the center of the star. The central temperature then rises and this in turn increases the rate of hydrogen burning until there is a new balance between heat produced at the center and the heat lost from the solar surface. Unlike ordinary fires, star-sized nuclear fires burn fiercer as the ash accumulates and sometimes even explode. Daisyworld is simplified, reduced if you like, in the following ways. The environment is reduced to a single variable, temperature, and the biota to a single species, daisies. If too cold, below 5°C, daisies will not grow; they do best at a temperature near 20°C. If the temperature exceeds 40°C, it will be too hot for the daisies, and they will wilt and die. The mean temperature of the planet is a simple balance between the heat received from the star and the heat lost to the cold depths of space in the form of long-wave infrared radiation. On the Earth, this heat balance is complicated by the effects of clouds and of gases such as carbon dioxide. The sunlight may be reflected back to space by the clouds before it can reach and warm the surface. On the other hand, the heat loss from the warm surface may be lessened because clouds and molecules of carbon dioxide reflect it back to the surface. Daisyworld is assumed to have a constant amount of carbon dioxide, enough for daisies to grow but not so much as to complicate the climate. Similarly, there are no clouds in the daytime to mar the simplicity of the model, and all rain falls during the night. The mean temperature of Daisyworld is, therefore, simply determined by the average shade of color of the planet, or as astronomers call it, the albedo. If the planet is a dark shade, low albedo, it absorbs more heat from the sunlight and the surface is warmed. If light in color, like fallen snow, then 70 or 80 percent of the sunlight may be reflected back to space. Such a surface is cold when compared with a dark surface under comparable solar illumination. Albedos range from 0 (wholly black) to 1 (wholly white). The bare ground of Daisyworld is usually taken to have an albedo of 0.4 so that it absorbs 40 percent of the sunlight that falls upon it. Daisies range in shade of color from dark (with an albedo of 0.2) to light (with an albedo of 0.7).
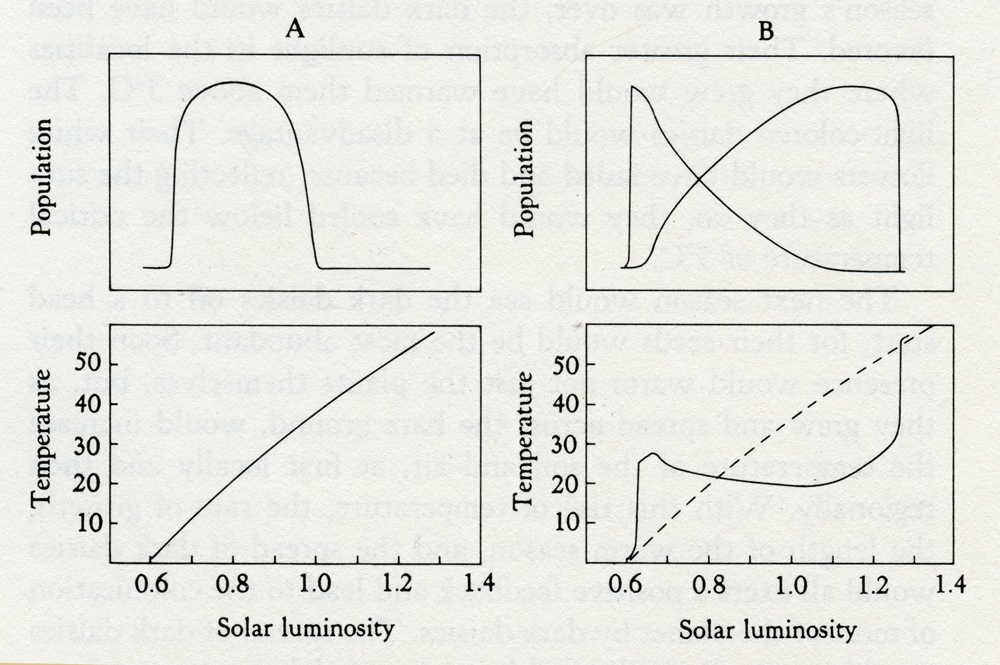
Imagine a time in the distant past of Daisyworld. The star that warms it was less luminous, so that only in the equatorial region was the mean temperature of bare ground warm enough, 5°C, for growth. Here daisy seeds would slowly germinate and flower. Let us assume that in the first crop multicolored, light, and dark species were equally represented. Even before the first season's growth was over, the dark daisies would have been favored. Their greater absorption of sunlight in the localities where they grew would have warmed them above 5°C. The light-colored daisies would be at a disadvantage. Their white flowers would have faded and died because, reflecting the sunlight as they do, they would have cooled below the critical temperature of 5°C. The next season would see the dark daisies off to a head start, for their seeds would be the most abundant. Soon their presence would warm not just the plants themselves, but, as they grew and spread across the bare ground, would increase the temperature of the soil and air, at first locally and then regionally. With this rise of temperature, the rate of growth, the length of the warm season, and the spread of dark daisies would all exert a positive feedback and lead to the colonization of most of the planet by dark daisies. The spread of dark daisies would eventually be limited by a rise of global temperature to levels above the optimum for growth. Any further spread of dark daisies would lead to a decline in seed production. In addition, when the global temperature is high, white daisies will grow and spread in competition with the dark ones. The growth and spread of white daisies is favored then because of their natural ability to keep cool. As the star that shines on Daisyworld grows older and hotter, the proportion of dark to light daisies changes until, finally, the heat flux is so great that even the whitest daisy crop cannot keep enough of the planet below the critical 40°C upper limit for growth. At this time flower power is not enough. The planet becomes barren again, and so hot that there is no way for daisy life to start again. It is easy to make a numerical model of Daisyworld simple enough to run on a personal computer. Daisy populations are modeled by differential equations borrowed from theoretical ecology (Carter and Prince, 1981). The mean temperature of the planet is calculated directly from the balance of the heat it receives from its star and the heat it loses by radiation to the cold depths of space. Figure 2.1 shows the evolution of the temperature and the growth of daisies during the progressive increase in heat flux from its star according to the conventional wisdom of physics and biology, and according to geophysiology.
Solar luminosity Solar luminosity 2.1 Models of the evolution of Daisyworld according to conventional wisdom (A) and to geophysiology (B). The upper panels illustrate daisy populations in arbitrary units; the lower panels, temperatures in degrees Celsius. Going from left to right along the horizontal axis, the star's luminosity increases from 60 to 140 percent of that of our own Sun. A illustrates how the physicists and the biologists in complete isolation calculate their view of the evolution of the planet. According to this conventional wisdom, the daisies can only respond or adapt to changes in temperature. When it becomes too hot for comfort, they will die. But in the Gaian Daisyworld (B), the ecosystem can respond by the competitive growth of the dark and light daisies, and regulate the temperature over a wide range of solar luminosity. The dashed line in the lower panel in B shows how the temperature would rise on a lifeless Daisyworld. When I first tried the Daisyworld model I was surprised and delighted by the strong regulation of planetary temperature that came from the simple competitive growth of plants with dark and light shades. I did not invent these models because I thought that daisies, or any other dark- and light-colored plants, regulate the Earth's temperature by changing the balance between the heat received from the Sun and that lost to space. I had designed them to answer the criticism of Ford Doolittle and Richard Dawkins that Gaia was teleological. In Daisyworld, one property of the global environment, temperature, was shown to be regulated effectively, over a wide range of solar luminosity, by an imaginary planetary biota without invoking foresight or planning. This is a definitive rebuttal of the accusation that the Gaia hypothesis is teleological, and so far it remains unchallenged. So what is Gaia? If the real world we inhabit is self-regulating in the manner of Daisyworld, and if the climate and environment we enjoy and freely exploit is a consequence of an automatic, but not purposeful, goal-seeking system, then Gaia is the largest manifestation of life. The tightly coupled system of life and its environment, Gaia, includes: 1. Living organisms that grow vigorously, exploiting any environmental opportunities that open. 2. Organisms that are subject to the rules of Darwinian natural selection: the species of organisms that leave the most progeny survive. 3. Organisms that affect their physical and chemical environment. Thus animals change the atmosphere by breathing: taking in oxygen and letting out carbon dioxide. Plants and algae do the reverse. In numerous other ways, all forms of life incessantly modify the physical and chemical environment. 4. The existence of constraints or bounds that establish the limits of life. It can be too hot or too cold; there is a comfortable warmth in between, the preferred state. It can be too acid or too alkaline; neutrality is preferred. Almost all chemicals have a range of concentrations tolerated or needed by life. For many elements, such as iodine, selenium, and iron, too much is a poison, too little causes starvation. Pure uncontaminated water will support little; but neither will the saturated brine of the Dead Sea. Few scientists would object to any of these four conditions, either singly or taken as a group. When they are taken together as a tightly coupled ensemble, they seem to form a recipe for a Gaian system. The ensemble is a fruitful source of models of self-regulating systems like Daisyworld. The fourth condition, which sets the physical and chemical bounds of life, I find the most interesting, unexpected, and full of insight. One has only to think of the social analogue of the family or community that exists with firm but reasonable bounds in comparison with one in which the limits of behavior are ill-defined. Stability and well-defined bounds seem to go together. Physicists are agreed that life is an open system. But like one of those Russian dolls which enclose a series of smaller and still smaller dolls, life exists within a set of boundaries. The outer boundary is the Earth's atmospheric edge to space. Within the planetary boundary, entities diminish but grow ever more intense as the inward progression goes from Gaia to ecosystems, to plants and animals, to cells and to DNA. The boundary of the planet then circumscribes a living organism, Gaia, a system made up of all the living things and their environment. There is no clear distinction anywhere on the Earth's surface between living and nonliving matter. There is merely a hierarchy of intensity going from the "material" environment of the rocks and the atmosphere to the living cells. But at great depths below the surface, the effects of life's presence fade. It may be that the core of our planet is unchanged as a result of life; but it would be unwise to assume it. In exploring the question, "What is life?" we have made some progress. By looking at life through Gaia's telescope, we see it as a planetary-scale phenomenon with a cosmological life span. Gaia as the largest manifestation of life differs from other living organisms of Earth in the way that you or I differ from our population of living cells. At some time early in the Earth's history before life existed, the solid Earth, the atmosphere, and oceans were still evolving by the laws of physics and chemistry alone. It was careering, downhill, to the lifeless steady state of a planet almost at equilibrium. Briefly, in its headlong flight through the ranges of chemical and physical states, it entered a stage favorable for life. At some special time in that stage, the newly formed living cells grew until their presence so affected the Earth's environment as to halt the headlong dive towards equilibrium. At that instant the living things, the rocks, the air, and the oceans merged to form the new entity, Gaia. Just as when the sperm merges with the egg, new life was conceived. The quest to define life might be compared with assembling a jigsaw puzzle, a puzzle where a landscape scene is cut into a thousand small interlocking pieces and the pieces scrambled. Classification is needed to put it together again. The blue sky is easy to separate from the brown earth and green trees. Skilled solvers of the jigsaw puzzle know that a key step is to find and connect the straight-sided pieces that define the edge, the boundary of the scene. The discovery that the outer reaches of the atmosphere are a part of planetary life in a like manner has defined the edge of our puzzle picture of the Earth. Once the edge is completely assembled, at least the size of the picture is known and the placing of the inner groupings made easier. Gaia is no static picture. She is forever changing as life and the Earth evolve together, but in our brief life span she keeps still long enough for us to begin to understand and see how fair she is.
|